
La reproduction de l’huître creuse fait intervenir une succession d’étapes, de la gamétogenèse à la fixation en passant par la phase larvaire, soumises à de nombreux aléas.
Le cycle de reproduction
La sexualité des huîtres a très tôt été décrite comme reposant sur un hermaphrodisme successif : elles peuvent être mâles et femelles alternativement mais la présence simultanée des deux sexes chez un même individu est rare (Amemiya, 1929). Toutefois, la détermination du sexe est un phénomène complexe qui semble régulé par de nombreux paramètres extérieurs (Lango Reynoso, 1999). Dans les premières études sur le sujet, toutes les possibilités de changements ont été observés lors de suivis individuels sur plusieurs années (Amemiya, 1929 ; Needler, 1942) avec cependant toujours une proportion importante d’huîtres qui restent mâles. C’est ce qui à conduit Coe (1934) à différencier, pour C. virginica, les “vrais mâles” des mâles hermaphrodites. La seconde observation générale consiste en la baisse du sexe-ratio mâle/femelle au fil du temps au sein d’une même classe d’âge vers une valeur de 1 (Amemiya, 1929 ; Buroker, 1983). De ces premières constatations, deux pistes de recherche ont été poursuivies en s’attachant, soit à montrer l’influence des facteurs environnementaux sur la détermination du sexe, soit à proposer un modèle génétique de détermination du sexe des huîtres creuses.
Pourtant, aujourd’hui encore les conditions qui conduisent une huîtres à être mâle ou femelle restent un mystère.
-

Cycle de reproduction de l’huître creuse, Crassotrea gigas.
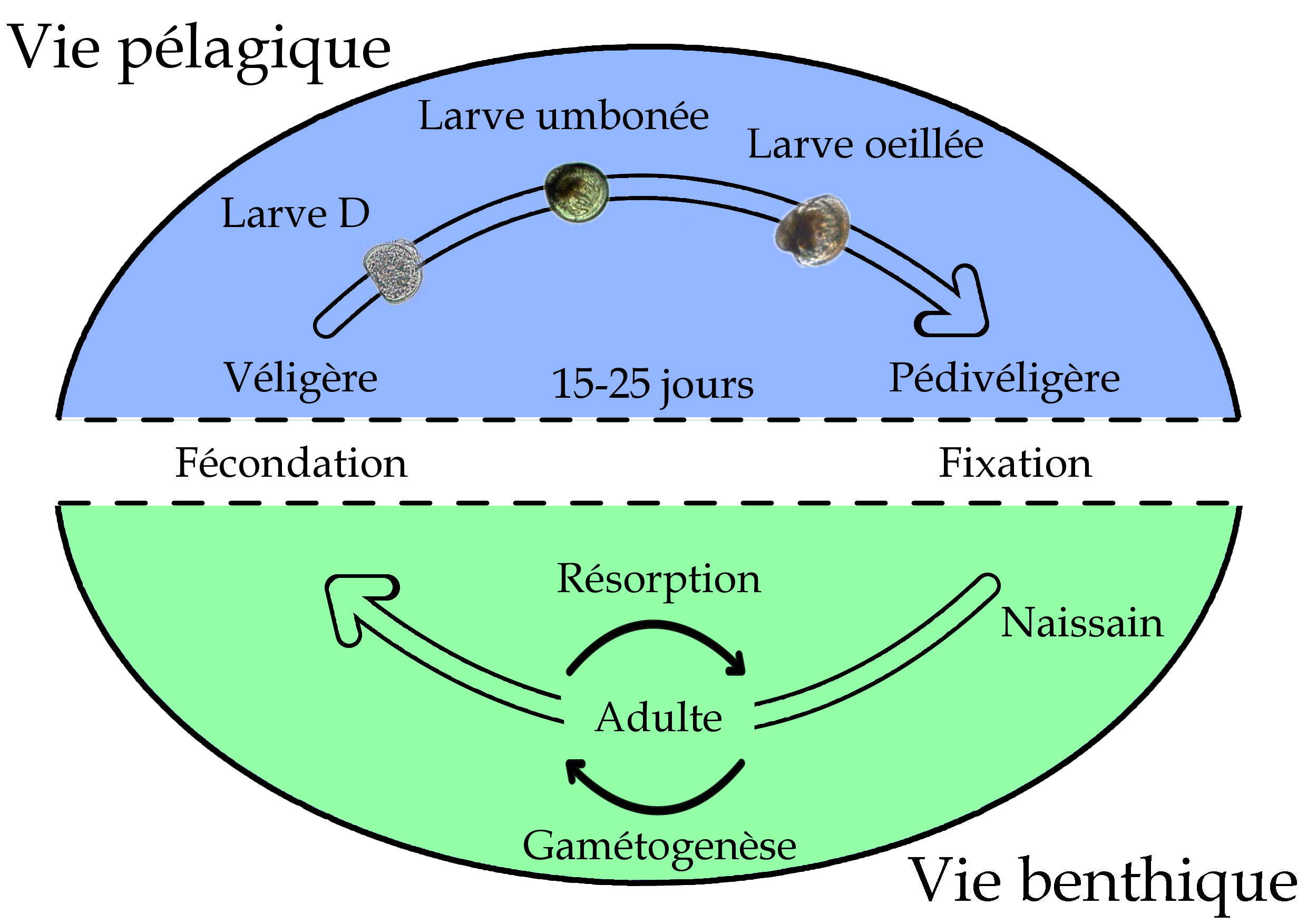
Le cycle de reproduction de l’huître creuse est composé de deux phases bien distinctes : une phase de maturation saisonnière des produits sexuels, la gamétogenèse, et une phase de vie larvaire (figure ci-dessus). Les périodes de transition entre ces phases sont brèves, il s’agit de la fécondation et de la fixation. Pour le bon succès de la reproduction, il est nécessaire que l’ensemble de ces étapes se passent sans accrocs.
La gamétogenèse
Au cours de la première phase, les géniteurs adultes sessiles maturent un grand nombre de gamètes. Cette maturation commence à la fin de l’hiver ce qui, sur les côtes françaises, correspond généralement à des températures de l’eau de 8 à 11°C (Le Dantec, 1968 ; Pouvreau et Le Pennec, 2006). Elle se poursuit ensuite jusque vers le mois de juillet.
La fécondation
A cette période, l’huître entre dans une période d’instabilité où des conditions favorables pourront déclencher l’émission des gamètes. L’émission des gamètes s’effectue donc principalement au cours des mois de juillet ou d’août. La fécondation est externe : la rencontre entre l’ovule et le spermatozoïde a lieu au sein de la colonne d’eau.
L’émission des gamètes chez l’huître creuse se traduit chez la femelle par de violents mouvements valvaires. Ceci rend la ponte de la femelle détectable au moyen d’un enregistrement de l’activité valvaire (Nelson, 1928 ;Galtsoff, 1938 ; His, 1970, 1975). Chez le mâle, c’est une importante action des cils qui propulse les spermatozoïdes à l’extérieur. Cette émission de gamètes peut être partielle ou totale en fonction de la quantité de gamètes et des conditions du milieu. Sa durée peut aller de quelques minutes à plus d’une heure. Elle a lieu préférentiellement durant le flot de la marée (Nelson, 1928 ; Carriker, 1951). Il existe généralement plusieurs émissions durant l’été.
L’importance de l’émission et sa synchronicité sont des facteurs de succès de la cohorte larvaire qui en résulte (His, 1975). Des conditions du milieu favorables, température supérieure à 20°C et nourriture abondante donnent lieu à des émissions de gamètes importantes et bien synchronisées tandis qu’un milieu défavorable provoquera des émissions diffuses (His, 1991).
La vie larvaire
A partir de ce moment débute la vie larvaire de l’huître creuse. La larve commence sa vie au stade de larve trochophore avant de rapidement devenir une larve véligère, au bout de 24h environ. A ce stade, elle mesure seulement 60 µm et vit encore de ses réserves énergétiques. Elle devient strictement planctotrophe, c’est à dire qu’elle se nourrit du phytoplancton, au bout de 5 jours environ (Rico-Villa et al., 2010, p. 87). La larve véligère dispose d’un velum, une sorte de voile cilié, qui lui sert à se déplacer et à capturer ses proies. La coquille formée de deux valves, ou prodissoconque, est déjà présente. La durée de la phase larvaire varie entre 15 et 25 jours sur la côte atlantique, en fonction de la température et de la nourriture disponible.
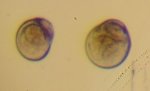
Larves d’huître creuse, Crassostrea gigas, au stade “petites évoluées”.
A la fin de la vie larvaire, lorsqu’elle atteint les 300 µm, la larve se munit d’un pied, elle devient pédivéligère. Cet organe va lui permettre de ramper pour choisir le substrat sur lequel elle va se fixer. C’est aussi cet organe qui sécrétera le ciment pour la fixation. Une fois fixée, la larve commence sa métamorphose et le développement des organes de l’adulte (Marteil, 1976).
La fixation
Voici comment Marteil (1976) décrit l’étape émouvante de la fixation de
la larve :
Quand arrive le moment de la fixation, la larve nage grâce à son velum, à la recherche d’un support solide, libre de vase ou de limon. Elle y fixe son pied et commence à ramper, le vélum rétracté, explorant ainsi la surface atteinte, se déplaçant valves en avant, allant et rampant “comme un chien préparant sa niche”
[. . .] Lorsqu’elle est prête à se fixer, la charnière surélevée, se balançant d’arrière en avant, et d’un côté à l’autre, elle expulse le contenu de la glande byssogène “comme d’un tube de seccotine” (Yonge, 1960), se tourne aussitôt sur la valve gauche qui s’applique à la goutte de ciment qui va durcir en quelques minutes et la maintiendra attachée. C’est donc par le bord supérieur de la valve gauche qu’elle se fixe et non par la charnière comme on le croit parfois. Une fois le ciment émis, la larve ne pourra plus se fixer s’il arrivait qu’elle soit détachée du support.
La larve, si elle peut explorer plusieurs substrats différents, ne peut se fixer qu’une seule fois. La présence d’un substrat adaptée à la fixation conditionne ainsi naturellement le captage. Un comportement grégaire des larves a été montré chez C. virginica, les larves se fixent préférentiellement sur les substrats où des huîtres sont déjà présentes (Hidu, 1969). Des éléments chimiques semblent être responsables de ce phénomène dans le milieu naturel (Crisp, 1967). Ainsi, l’epinephrine est connue pour induire la métamorphose chez C. gigas tandis que la L-3,4-dihydroxyphenylalanine (L-DOPA) induit un comportement de fixation (Coon et al., 1990). A l’aide de ces stimulants chimiques, il a été montré une déconnection entre le comportement de fixation et la métamorphose et plus particulièrement la possibilité pour la larve de C. gigas de retarder la métamorphose, tout en conservant sa compétence (Coon et al., 1990).
Cette petite huître, tout nouvellement fixée, va pouvoir elle aussi produire des gamètes pour se reproduire si elle échappe à ses prédateurs.
Cette description de la reproduction de l’huître creuse est en partie issue de mes travaux de thèse, sur la variabilité de la reproduction de l’huître creuse. Cette thèse est téléchargeable ici.
Références citées :
Amemiya, I. (1929), ’On the sex-change of the japanese Common Oyster, Ostrea gigas Thunberg’, Proceding of the Imperial Academy of Tokyo 5, 284—286.
Buroker, N. E. (1983), ’Sexuality with respect to shell length and group size in the japanese oyster Crassostrea gigas’, Malacologia 23(2), 271—279.
Carriker, M. R. (1951), ’Ecological observations on the distribution of oyster larvae in New Jersey estuaries’, Ecological Monographs 21(1), 19—38.
Coe, W. R. (1934), ’Alternation of sexuality in oysters’, American Naturalist 68(716), 236—251.
Coon, S. L. ; Fitt, W. K. & Bonar, D. B. (1990), ’Competence and delay of metamorphosis in the Pacific oyster Crassostrea gigas’, Marine Biology 106(3), 379—387.
Crisp, D. J. (1967), ’Chemical factors inducing settlement in Crassostrea virginica (Gmelin)’, Journal of Animal Ecology 36, 329—335.
Galtsoff, P. S. (1938), ’Physiology of reproduction of Ostrea virginica : I. spawning reactions of the female and male’, The Biological Bulletin 74(3), 461—486.
Hidu, H. (1969), ’Gregarious setting in the american oyster Crassostrea virginica Gmelin’, Chesapeake science 10(2), 86—92.
His, E. (1991), ’Biologie et écotoxicologie des véligères de Crassostrea gigas (Thunberg) dand le bassin d’Arcachon’, Technical report, Universitй Bordeaux I.
His, E. (1975), ’La détection des pontes dans le milieu naturel : application de l’ostréographie à l’étude de la reproduction des huîtres’, haliotis 5, 206—215.
His, E. (1970), ’L’émission des gamètes chez l’huître portugaise (Crassostrea angulata LMK)’, Revue des Travaux de l’Institut des Pêches Maritimes 34(1), 17—22.
Reynoso, F. L. (1999), ’Détermination de la sexualité chez l’huître Crassostrea gigas (Thunberg, 1793)’, PhD thesis, Universitй de Bretagne Occidentale.
Dantec, J. L. (1968), ’Ecologie et reproduction de l’huitre portugaise (Crassostrea angulata Lamarck) dans le bassin d’Arcachon et sur la rive gauche de la Gironde’, Revue des Travaux de l’Institut des Pêches Maritimes 32(3), 237—362.
Marteil, L. (1976), ’La conchyliculture française – Biologie de l’huître et de la moule’, Revue des Travaux de l’Institut des Pêches Maritimes 40(2), 149—346.
Needler, A. B. (1942), ’Sex reversal in individual oysters’, J. Fish. Res. Can. 5(4), 361—364.
Nelson, T. C. (1928), ’Relation of Spawning of the Oyster to Temperature’, Ecology 9(2), 145-154.
Pouvreau, S. & Le Pennec, M. (2006), ’Ecophysiologie de la reproduction chez l’huître creuse, Crassostrea gigas’, Technical report, IFREMER.
Rico-Villa, B. ; Bernard, I. ; Robert, R. & Pouvreau, S. (2010), ’A Dynamic Energy Budget (DEB) growth model for Pacific oyster larvae, Crassostrea gigas’, Aquaculture 305(1-4), 84—94.